Биоэнергетика клеток и ее связь с подвижностью
микроорганизмовУчебные материалы по биологии / Идентификация микроводорослей Euglena glacilis и анализ их чувствительности к ингибирующим веществам / Биоэнергетика клеток и ее связь с подвижностью
микроорганизмовСтраница 3
Перенос электронов (дыхание)→ ![]() →АТФ
→АТФ
Сопряжение окисления и фосфорилирования через![]() позволяет объяснить, почему О.ф., в отличие от гликолитического ("субстратного") фосфорилирования, протекающего в растворе, возможно лишь в замкнутых мембранных структурах, а также почему все воздействия, снижающие электрическое сопротивление и увеличивающие протонную проводимость мембраны, подавляют ("разобщают") О.ф. Энергия
позволяет объяснить, почему О.ф., в отличие от гликолитического ("субстратного") фосфорилирования, протекающего в растворе, возможно лишь в замкнутых мембранных структурах, а также почему все воздействия, снижающие электрическое сопротивление и увеличивающие протонную проводимость мембраны, подавляют ("разобщают") О.ф. Энергия ![]() , помимо синтеза АТФ, может непосредственно использоваться клеткой для других целей - транспорта метаболитов, движения (у бактерий), восстановления никотинамидных коферментов и др.
, помимо синтеза АТФ, может непосредственно использоваться клеткой для других целей - транспорта метаболитов, движения (у бактерий), восстановления никотинамидных коферментов и др.
В дыхательной цепи имеется несколько участков, которые характеризуются значит. перепадом окислительно-восстановительного потенциала ∆Е и сопряжены с запасанием энергии (генерацией ![]() ). Таких участков, называющих пунктами или точками сопряжения, обычно три: убихинон-редуктазное звено (∆Е~0,35-0,4 В), убихинол: цитохром-c-редуктазное звено (∆Е~0,25 В) и цитохром-с-оксидазный комплекс (∆Е~0,6 В) - пункты сопряжения 1, 2 и 3 соотв. В соответствии с рисунком 9. Каждый из пунктов сопряжения дыхательной цепи может быть выделен из мембраны в виде индивидуального ферментного комплекса, обладающего окислительно-восстановительной активностью. Такой комплекс, встроенный в фосфолипидную мембрану, способен функционировать как протонный насос [14].
). Таких участков, называющих пунктами или точками сопряжения, обычно три: убихинон-редуктазное звено (∆Е~0,35-0,4 В), убихинол: цитохром-c-редуктазное звено (∆Е~0,25 В) и цитохром-с-оксидазный комплекс (∆Е~0,6 В) - пункты сопряжения 1, 2 и 3 соотв. В соответствии с рисунком 9. Каждый из пунктов сопряжения дыхательной цепи может быть выделен из мембраны в виде индивидуального ферментного комплекса, обладающего окислительно-восстановительной активностью. Такой комплекс, встроенный в фосфолипидную мембрану, способен функционировать как протонный насос [14].
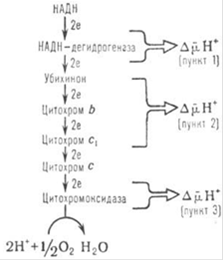
Рисунок 9 - Упрощенная схема расположения пунктов сопряжения в цепи дыхательных ферментов; НАДН - восстановленная форма кофермента никотинамидадениндинуклеотида.
Обычно для характеристики эффективности О.ф. используют величины Н+/2е или q/2e, указывающие сколько протонов (либо электрических зарядов) переносится через мембрану при транспорте пары электронов через данный участок дыхательной цепи, а также отношение Н+/АТФ, показывающее, сколько протонов нужно перенести снаружи внутрь митохондрий через АТФ-синтетазу для синтеза 1 молекулы АТФ. Величина q/2e составляет для пунктов сопряжения 1, 2 и 3 соответственно 3-4, 2 и 4. Величина Н+/АТФ при синтезе АТФ внутри митохондрий равна 2; однако еще один Н+ может тратиться на вынос синтезированного АТФ 4 из матрикса в цитоплазму переносчиком адениновых нуклеотидов в обмен на АДФ 3 . Поэтому кажущаяся величина Н+/АТФнаружн равна 3.
В организме О.ф. подавляется многими токсичными веществами, которые по месту их действия можно разделить на три группы:
) ингибиторы дыхательной цепи, или т. наз. дыхательные яды;
) ингибиторы АТФ-синтетазы. Наиболее распространенные ингибиторы этого класса, употребляемые в лабораторных исследованиях, - антибиотик олигомицин и модификатор карбоксильных групп белка дициклогексилкарбодиимид;
) так называемые разобщители О.ф. Они не подавляют ни перенос электронов, ни собственно фосфорилирование АДФ, но обладают способностью уменьшать величину ![]() на мембране, благодаря чему нарушается энергетическое сопряжение между дыханием и синтезом АТФ. Разобщающее действие проявляет большое число соединений самой разнообразной химической структуры. Классические разобщители - вещества, обладающие слабыми кислотными свойствами, способные проникать через мембрану как в ионизованной (депротонированной), так и в нейтральной (протонированной) формах. К таким веществам относят, напр., 1-(2-дицианометилен)гидразино-4-трифтор-метоксибензол, или карбонилцианид-n-трифторметокси-фенилгидразон, и 2,4-динитрофенол (соотв. формулы I и II; показаны протонированные и депротонированные формы) в соответствии с рисунком 10.
на мембране, благодаря чему нарушается энергетическое сопряжение между дыханием и синтезом АТФ. Разобщающее действие проявляет большое число соединений самой разнообразной химической структуры. Классические разобщители - вещества, обладающие слабыми кислотными свойствами, способные проникать через мембрану как в ионизованной (депротонированной), так и в нейтральной (протонированной) формах. К таким веществам относят, напр., 1-(2-дицианометилен)гидразино-4-трифтор-метоксибензол, или карбонилцианид-n-трифторметокси-фенилгидразон, и 2,4-динитрофенол (соотв. формулы I и II; показаны протонированные и депротонированные формы) в соответствии с рисунком 10.
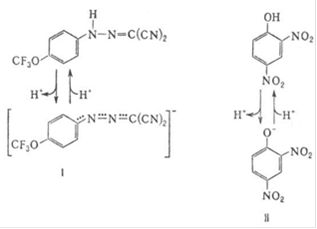
Рисунок 10 - Разобщителеи окислительного фосфолирования (2,4 - динитрофенол)
Двигаясь через мембрану в электрическое поле в ионизованной форме, разобщитель уменьшает ∆Ѱ; возвращаясь обратно в протонированное состоянии, разобщитель понижает ∆рН в соответствии с рисунком 11. Таким образом, такой "челночный" тип действия разобщителя приводит к уменьшению![]() .
.
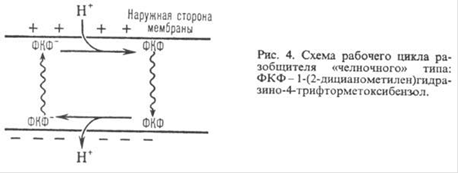
Рисунок 11 - Схема рабочего цикла разобщителя "челночного" типа: ФКФ - 1-(2-дицианометилен) гидразино-4-трифтометоксибензол
Разобщающим действием обладают также ионофоры (напр., грамицидин), повышающие электропроводность мембраны в результате образования ионных каналов или вещества, разрушающие мембрану (напр., детергенты).
О.ф. открыто В. А. Энгельгардтом в 1930 при работе с эритроцитами птиц. В 1939 В. А. Белицер и Е. Т. Цыбакова показали, что О.ф. сопряжено с переносом электронов в процессе дыхания; к такому же заключению несколько позднее пришел Г. М. Калькар.
Смотрите также
Структура и транспорт андрогенов
ВСТУПЛЕНИЕ
Реферат
посвящён обзорному исследованию мужских половых гормонов - андрогенов.
Актуальность
выбранной темы заключается в том, что уровень репродуктивного здоровья в мире ...
Строение, свойства и функции белков
...
Видовое разнообразие и морфометрические показатели бесхвостых земноводных Гомельского района
ВВЕДЕНИЕ
Земноводные
- низшие наземные позвоночные, сохраняющие связь с водой. Амфибии - одна из
древнейших групп животных. За 200 млн. лет существования возникло множество
форм земн ...