Эпигенетическая регуляция функций гонадотрофовУчебные материалы по биологии / Лютеинизирующий и фолликулостимулирующий гормоны. Физиологическая роль и механизмы регуляции / Эпигенетическая регуляция функций гонадотрофовСтраница 1
гонадотропин репродуктивный яичник гормон
В последнее время были достигнуты значительные успехи в понимании того, как изменения структуры хроматина провоцируют угнетение транскрипции каких-либо генов. Определённые трансформирующие ферменты обеспечивают посттрансляционную модификацию «хвостов» гистонов, например ацетилирование или метилирование. Эти модификации выражаются изменением генной экспрессии. Совсем недавно была выявлена связь между модификациями хвостов гистонов и ключевыми процессами в гипофизарных гонадотрофах, включая интересующие нас процессы транскрипции генов β-субъединиц гонадотропинов и влияние ГнРГ на эти процессы.
Ацетилирование и деацетилирование гистонов, ассоциированных с участками - промоторами генов β-субъединиц гонадотропинов, играет важнейшую роль в активации и супрессии этих генов. Этот динамический процесс связан с вовлечением и активацией двух групп трансформирующих ферментов: гистоновых ацетилтрансфераз (HATs) и гистоновых деацетилаз (HDACs). HATs ацетилируют лизиновые остатки хвостов гистонов, и это рассматривается как процесс, вызывающий локальную деконденсацию хроматина. Деконденсация, в свою очередь, повышает доступность генных промоторных участков для транскрипционных факторов и увеличивает возможность образования комплекса «РНК-полимераза - промотор» и инициации транскрипции. Напротив, ферменты группы HDAC деацетилируют гистоны; при этом их «хвосты» приобретают положительный заряд, и это приводит к возрастанию компактизации хроматина (обусловленной электростатическими взаимодействиями между отрицательно заряженной ДНК и положительно заряженными гистонами). Некоторые из последних исследований приводят доказательства влияния ГнРГ на стимуляцию генов β-ЛГ и β-ФСГ субъединиц, опосредованного вышеописанной модификацией гистонов.
Вследствие отсутствия экспрессии β-субъединиц ЛГ и ФСГ и, соответственно, отсутствия секреции гонадотропинов, считается, что αT3-1-клеточная линия гонадотрофов представляет собой совокупность незрелых гонадотрофов. Согласно результатам исследований, различные ферменты группы HDAC в клетках этой линии подавляют транскрипцию генов β-субъединиц гонадотропинов практически до нулевого уровня, «оккупируя» их промоторы, и вызывают т.н. «транскрипционную тишину». Воздействие ГнРГ на тип клеток вышеозначенной линии (αT3-1) вызывает экспорт из ядра HDAC класса IIА, и это делает возможным предположение, что ГнРГ потенциально может менять состояние гистонов (т.е. присутствие или отсутствие ацетильных групп в их составе) и индуцировать генную экспрессию, элиминируя факторы, подавляющие транскрипцию. Этот факт доказывает, что ГнРГ модулирует активность гистонмодифицирующих ферментов в гонадотрофах. Более того, было обнаружено что с промоторами генов β-субъединиц ЛГ и ФСГ связаны разные типы HDAC, что позволяет сделать предположение о механизме, посредством которого осуществляется дифференциальная регуляция ЛГ и ФСГ в рамках клеток одной линии.
Исследования более зрелых гонадотрофов LβT2-клеточной линии, которые экспрессируют преимущественно мРНК βЛГ и, в меньшей степени, мРНК βФСГ, показали, что ферменты HAT опосредуют ГнРГ-стимуляцию обоих видов β-субъединиц гонадотропинов. HAT p300 ассоциирован с геном β-субъединицы ЛГ, и ГнРГ это связывание усиливает. В то же время, p300, действуя совместно с транскрипционными факторами SF1, Sp1 и Egr-1, играет важнейшую роль в усилении транскрипции β-субъедницы ЛГ. Было показано, что паралог p300 в LβT2-клетках, CREB-связывающий белок (СВР), ассоциирован с промотором гена β-субъединицы ФСГ. ГнРГ может усиливать эту связь путём взаимодействий с промотор-связанным фосфорилированным CREB, и стимулировать таким образом транскрипцию β-субъединицы ФСГ. (Рис.3) Дезактивация каким-либо образом р300 приводит к снижению транскрипции обоих гонадотропных β-субъединиц, и в регуляцию транскрипции включается НАТ.
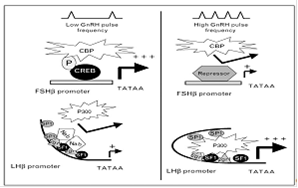
Рис.6. [5]
Итак, изучение αT3-1 и LβT2-клеточных линий подтверждает, что модификация гистонов посредством динамического ГнРГ-контроля HDAC и HAT может повлиять на присутствие/отсутствие ацетильных групп на хвостах гистонов, ассоциированных с генами-регуляторами гонадотрофной транскрипции в целом и транскрипции β-субъединиц гонадотропинов в частности. Так или иначе, нет исследований, показывающих корреляцию между ассоциацией HDAC и HAT с промоторами генов гонадотропинов и присутствием/отсутствием ацетильных групп на гистонах. Будущие исследования, сосредоточенные в этой области, будут способствовать дальнейшему пониманию относительной важности эпигенетических модификаций в регуляции гонадотропинов, в особенности в ответ на пульсирующий выброс ГнРГ. Кроме того, необходимо изучить другие модификации гистонов (метилирование, фосфорилирование) с точки зрения регуляции ЛГ и ФСГ.
Смотрите также
Окружающая среда и ее компоненты
Введение
Все
живые организмы, населяющие нашу планету, существуют не сами по себе, они
зависят от окружающей среды и испытывают на себе ее воздействия. Это точно
согласованный компле ...
Медоносные растения окрестностей селения Миглакаси Сергокалинского района
Введение
Растительный
мир селении Миглакаси очень разнообразен. При изучении растительного покрова
сел. Миглакаси раскрываются черты его замечательного богатства и своеобразия.
Именн ...
Систематика растений
Лекция
№1
...