Метаболические пути синтезаУчебные материалы по биологии / Генетика и биохимия микробного синтеза полигидроксиалканоатов / Метаболические пути синтезаСтраница 1
На примере наиболее изученного из ПГА - полимера β-оксимасляной кислоты установлено, что пути его синтеза практически одинаковы у различных микроорганизмов (Alkaligenes, Azotobakter, Pseudomonas).
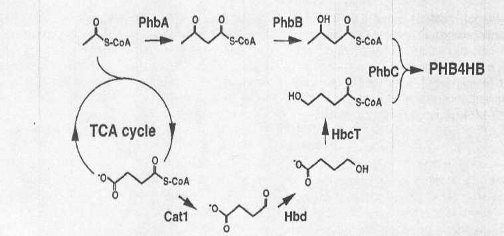
Рис.2. Биосинтез P (3HB-4HB) в рекомбинантном E. coli при использовании гетерологичных генов от Clostridlum kluyveri.
HB мономер в синтезе P (3HB-4I IB) получен из сукцината. В сукцинате преобразовывают 4HB-CoA ферменты, которые образовывают дугу закодированный генами от грамположительного, строго анаэробного C. kluyveri микроба.
В биосинтезе полигидроксиалканоатов ключевыми являются три фермента: β-кетотиолаза, ацетоацетил-КоА-редуктаза и ПГА-синтаза, которые кодируются соответственно генами phbA, phbB, phbC (Рис.3). Регуляция процесса синтеза ПГА может осуществляться на нескольких уровнях: на уровне экспрессии генов специфическими факторами среды (например, недостаток питательных элементов) или на уровне регуляции активности ферментов специфическими клеточными компонентами, являющимися их субстратами или ингибиторами. Возможен смешанный тип регуляции активности ферментов, участвующих в метаболизме ПГА. В самых первых работах, посвященных изучению механизма синтеза ПГА, было показано, что регуляция синтеза ПГА у R.eutropha осуществляется метаболитами на ферментативном уровне при ведущей роли внутриклеточной концентрации свободного коэнзима А, который ингибирует кетотиолазную реакцию у A.eutrophus, у Azotobacter beijerinckii, а также у Zoogloea ramigera. Установлено, что синтез ПГА стимулируется высокой внутриклеточной концентрацией НАДФН и высоким соотношением НАДФН/НАДФ.
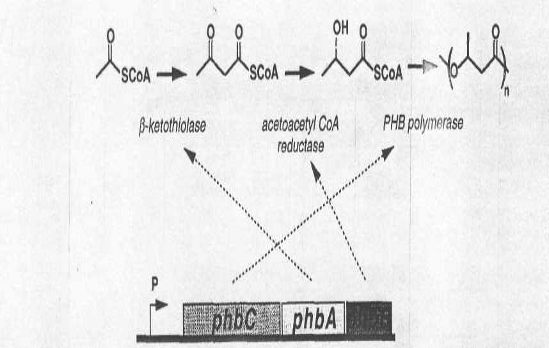
Рис.3. Схематичное изображение генов ферментов синтеза ПГА и продуктов их экспрессии.
β-кетотиолаза является первым ферментом синтеза полимера и катализирует конденсацию двух молекул ацетил-КоА, фермент также катализирует последнюю реакцию в процессе эндогенной деградации полимера. Таким образом, этот фермент играет ключевую роль в циклическом метаболизме полимера. Активность фермента не ингибируется физиологическими концентрациями метаболитов клетки, но реакция конденсации резко снижается при накоплении свободного КоА. Эффект ингибирования свободным КоА синтеза полимера является очень важным аспектом метаболической регуляции клетки, так как в условиях повышенных концентраций свободного КоА субстраты используются главным образом в биосинтетических реакциях, приводящих к синтезу белков, нуклеиновых кислот и других первичных макромолекул. При этом ацетил КоА используется в основном цикле трикарбоновых кислот (ЦТК), где на первом этапе цикла цитратсиназа высвобождает свободный КоА. В этих условиях внутриклеточная концентрация свободного кофермента значительно выше, чем в условиях несбалансированного роста. В результате высокий уровень КоА ингибирует активность β-кетотиолазы и в конечном итоге,- синтез полимера. О регуляторной роли ЦТК в направлении потоков ацетил КоА в клетке свидетельствуют данные, полученные на мутанте R.eutropha, лишенного изоцитратдегидрогеназной активности, который синтезирует со значительно большей скоростью, чем дикий штамм. Вместе с тем, показано, что активность первых двух ферментов ЦТК ингибируется повышенными концентрациями восстановленных никотинамидных нуклеотидов при несбалансированном росте. Таким образом, синтез полимера, например, в R.eutropha начинается, когда ингибируются активности ферментов ЦТК (цитратсиназы и изоцитратдегидрогеназы). Это приводит к увеличению внутриклеточной концентрации ацетил-КоА, восстановленных пиридиннуклеотидов, снижению концентрации свободного кофермента А и окисленных форм нуклеотидов. Сходный контроль ферментативной активности на уровне метаболитов был подтверждён на рекомбинантном штамме E.coli, трансформированном генами синтеза полимера из R.eutropha.
На первых порах в R.eutropha было обнаружено две β-кетотиолазы. Уровень ферментативной активности кетотиолаз в клетках, выращенных в условиях лимитирования роста азотом или углеродом, одинаков (0,20 и 0,27 U/мг белка, соответственно). Это позволило авторам сделать вывод о конститутивной природе фермента. Было установлено, что две кетотиолазы отличаются субстратной специфичностью. Так β-кетотиолаза А активна с ацетоацетил-КоА и 3-кетопентаноил-КоА и является главным претендентом на участие в синтезе субстратов для ПГА. Субстратная специфичность β-кетотиолазы В оказалась более широкой, фермент активен с длинноцепочечными 3-кетоацильными производными КоА от 4 до 10 атомов углерода. Было высказано предположение, что эта кетотиолаза является ферментом β-окисления жирных кислот. Участие кетотиолазы В в синтезе субстратов для образования полимера было подтверждено на рекомбинантных штаммах Escherichia coli, дефектным по генам, ответственным за деградацию жирных кислот и транспорт пропионата, и содержащим оперон синтеза полимера из A.eutrophus без гена кетотиолазы (phaA). В дальнейшем из R.eutropha были клонированы три последовательности, кодирующие три β-кетотиолазы. Было установлено, что β-кетотиолаза, кодируемая phaA геном, является β-кетотиолазой А и высокоактивна с ацетоацетил-КоА. Вторая β-кетотиолаза, являющаяся продуктом bktB гена, отвечает преимущественно за синтез кетовалерил-КоА и в меньшей степени - кетогексаноил-КоА, а также, возможно, соответствует β-кетотиолазе В и участвует в β-окислении коротко- и среднецепочечных жирных кислот. Функция третьей кетотиолазы пока до конца не выяснена, но она также активна с ацетоацетил-КоА как субстратом. Участие β-кетотиолазы В в синтезе полимера было подтверждено в работе. Авторы установила, что в условиях блокирования β-окисления жирных кислот в клетках R.eutropha создается пул среднецепочечных 3-гидроксикислот, служащих субстратом для синтеза ПГА. В результате клетки накапливают гетерополимерные ПГА с включением гидроксигексаноата до 7-10 мол.%.
Смотрите также
Теория "естественного отбора" Ч. Дарвина
Введение
В 2001 году исполнилось 142 года со дня выхода в свет
“Происхождения видов” и торжества теории эволюции. Просвещенная общественность
настолько сжилась с представлениями Дарви ...
Виды и функции иммунитета. Пектиновое и анаэробное брожение клетчатки
...
Научные революции ХХ века
Введение
В
течение довольно длительного времени господствовало представление о том, что
развитие науки происходит путем постепенного, непрерывного накопления все новых
и новых научны ...