Эндодерма и центральный цилиндрУчебные материалы по биологии / Транспорт и распределение тяжелых металлов и поливалентных катионов в высших растениях / Эндодерма и центральный цилиндрСтраница 2
Выявленные различия распределения Ni и Sr в клетках перицикла определяют различия в накоплении и распределении этих металлов по всем остальным тканям центрального цилиндра. Поступление Ni из клеток перицикла в другие ткани центрального цилиндра ограничено. Причина подобного явления может заключаться в том, что транспорт Ni в центральном цилиндре происходит по симпласту, а большая часть плазмодесм в перицикле расположена в наружных тангентальных стенках клеток на границе с эндодермой и примерно столько же − в радиальных. Имея мощный “вход” Ni из клеток коры, клетки эндодермы и перицикла структурно имеют значительно меньший “выход”, аккумулируя, таким образом, Ni в протопластах. Конечно, подобное явление было бы невозможным, если бы ионы Ni могли, подобно Sr, проходить через плазмалемму клеток стели и транспортироваться по апопласту.
В отличие от Ni, содержание Sr в клеточных оболочках клеток перицикла и центрального цилиндра уже после двух суток инкубации было приблизительно одинаковым. Следовательно, функциональная роль перицикла как “коллектора” ионов не универсальна для всех элементов, а свойственна, по-видимому, только ионам, передвигающимся по симпласту.
В связи с проблемой гипераккумуляции, важно установить, играет ли перицикл роль коллектора у растений-гипераккумуляторов. Было показано, что даже при инкубации растений Thlaspi caerulescens в течение 1.5 мес. на питательном растворе с высокими концентрациями Ni(NO3)2 (300−400 мкМ) его накопления в перицикле не происходило. Таким образом, в корнях растений-гипераккумуляторов отсутствуют не только барьеры для транспорта Ni, но и, что, возможно, более важно, отсутствуют ткани-аккумуляторы и ткани-“коллекторы” и, как следствие, корневая система не играет барьерной роли, свойственной растениям-исключателям [].
Ультраструктура клеток перицикла, наряду с некоторыми признаками дифференцированных тканей, имеет черты, характерные для меристем. Можно говорить о двойственности функций перицикла. С одной стороны, это ткань, через которую вещества транспортируются к проводящим тканям, а с другой стороны, это меристематическая ткань, специфика которой заключается в способности давать начало боковым корням.
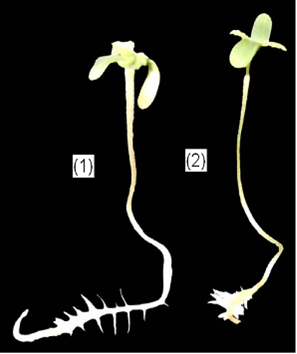
Рис. 7 Проростки конопли после 7 дней инкубации при действии Pb(NO3)2 в концентрации 10-4 М
-контроль, 2- Pb(NO3)2, 10-4 M [17].
Процесс развития боковых корней чрезвычайно устойчив к действию большинства тяжелых металлов. Даже при очень сильном ингибировании роста главного корня в растворах солей тяжелых металлов боковые корни возникают в то же время, что и у контрольных корней. Более того, многие авторы даже указывают на то, что под воздействием Cd и Pb наблюдалось образование более компактной и разветвленной корневой системы за счет развития дополнительных боковых корней (рис. 7) [6, 7, 17].
Однако причины устойчивости ветвления корней к воздействию тяжелых металлов могут быть различны. С одной стороны Cd и Pb не проникают в перицикл и, следовательно, не могут влиять на развитие боковых корней, с другой, металлы, проникая через эндодермальный барьер, могут впоследствии накапливаться в метаболически малоактивном компартменте - клеточной оболочке. Так происходит в случае сo Sr, что, наряду с его слабой токсичностью, и обуславливает нормальное заложение боковых корней в перицикле.
Уникальным в этом отношении является Ni, для которого клетки перицикла выполняют роль аккумулятора и “кольцевого коллектора”. Накапливаясь в протопласте и обладая высокой цитотоксичностью, Ni, единственный среди изученных металлов, ингибирует ветвление корня.
Ингибирующее действие Ni на образование боковых корней не наблюдалось у растений-гипераккумуляторов, что связано с низким содержанием Ni в клетках перицикла даже при высоких концентрациях соли Ni в растворе. Это может быть связано с тем, что перицикл гипераккумуляторов не играет роли аккумулятора и «коллектора» Ni и не препятствует его поступлению в ксилему.
Паренхимные клетки стелы включают в себя ксилемную паренхиму и паренхиму сердцевины, клетки которых отличаются по своей структуре. Клетки ксилемной паренхимы в корнях кукурузы имеют утолщенные клеточные оболочки, что также отличает их от паренхимы первичной коры. Поступая через эндодермальный барьер, уже через сутки после начала инкубации Sr выявлялся в апопласте клеток всех тканей стелы. Данное явление легко объяснить, если принять во внимание, что прочность связывания Sr с материалом клеточных оболочек невысока. Кроме того, значительная часть металла находится на поверхности клеточных оболочек или в периплазматическом пространстве. В совокупности это может определять высокую мобильность Sr, в результате чего модификация клеточных оболочек паренхимы стелы не ограничивает его передвижение в более глубокие ткани корня, в том числе в клетки сердцевины.
Смотрите также
Роль фауны в жизни леса
Введение
Лес - сложное образование природы, явление биологическое и
физическо - географическое, составная часть географического ландшафта и
биосферы нашей планеты.
Трудно себе пред ...
Человек как часть живого вещества
Введение
В 1916-м году, когда отечественный ученый В.И. Вернадский
ввел в науку представление о "живом веществе", это совершенно
изменило господствующее до того времени науч ...
Изучение ретровирусов
Введение
Вирус
(от лат. virus - яд) - простейшая форма жизни на нашей планете,
микроскопическая частица, представляющая собой молекулы нуклеиновых кислот (ДНК
или РНК), заключенные в ...