Материалы и методыУчебные материалы по биологии / Возможность использования украинских фамилий в качестве квазигенетических маркёров / Материалы и методыСтраница 2
![]()
![]() ,
,
где ![]()
![]() - тотальный инбридинг, определяемый соотношением случайного и неслучайного инбридинга; fn - неслучайный инбридинг, связанный с положительной или же отрицательной брачной ассортативностью (соответствует F[S); fr - случайный инбридинг, связанный с подразделенностью тотальной популяции, оценивает вклад j-той субпопуляции в дифференциацию тотального генофонда [3].
- тотальный инбридинг, определяемый соотношением случайного и неслучайного инбридинга; fn - неслучайный инбридинг, связанный с положительной или же отрицательной брачной ассортативностью (соответствует F[S); fr - случайный инбридинг, связанный с подразделенностью тотальной популяции, оценивает вклад j-той субпопуляции в дифференциацию тотального генофонда [3].
Для любой популяции (j) можно оценить величину случайного инбридинга по частотам фамилий следующим образом:
![]()
![]() = I/4,
= I/4,
где I - ожидаемая частота изонимных (однофамильных) браков в j-той популяции, т.е.
![]()
где ![]()
![]() - частота i-той фамилии в j-той субпопуляции, а коэффициент 1/4 учитывает передачу фамилий лишь по мужской линии [3].
- частота i-той фамилии в j-той субпопуляции, а коэффициент 1/4 учитывает передачу фамилий лишь по мужской линии [3].
. Расчёт частоты аллелей белковых локусов ![]()
![]() ,
, ![]()
![]() ,
, ![]()
![]() R, r.
R, r.
Закон Харди-Вайнберга гласит, что процесс наследственной преемственности сам по себе не ведет к изменению частот аллелей и (при случайном скрещивании) частот генотипов по определенному локусу. Более того, при случайном скрещивании равновесные частоты генотипов по данному локусу достигаются за одно поколение, если исходные частоты аллелей одинаковы у обоих полов. Равновесные частоты генотипов задаются произведениями частот соответствующих аллелей. Если имеются только два аллеля, А и а, с частотами р и q, то частоты трех возможных генотипов выражаются уравнением:
(p+q)2 = p2 + 2рq + q2
А а АА Аа аа,
где буквам во второй строке, обозначающим аллели и генотипы, соответствуют расположенные над ними частоты в первой строке.
Если имеются три аллеля, скажем А1, А2 и А3, с частотами р, q и r, то частоты генотипов определяются следующим образом:
(p + q + r)2 = р2 + q2 + r2 + 2pq+ 2рr + 2qr
А1 А2 А3 А1А1 А2А2 А3А3 А1А2 А1А3 А2А3
Аналогичный прием возведения в квадрат многочлена может быть использован для определения равновесных частот генотипов при любом числе аллелей. Заметим, что сумма всех частот аллелей, так же как и сумма всех частот генотипов, всегда должна быть равна 1. Если имеются только два аллеля с частотами р и q, то р + q = 1, и, следовательно, р2 + 2рq + q2 = (р + q)2 = 1; если же имеется три аллеля с частотами р, q и r, то р + q + r = 1, и, следовательно, также (р + q + r)2 =1 и т.д. [1].
. Расчёт генетических расстояний.
Для расчёта генетических расстояний между популяциями с использованием фамилий и сравнения их с генетическими расстояниями, рассчитанными по данным о частотах аллелей локусов ![]()
![]() ,
, ![]()
![]() ,
, ![]()
![]() R, r, использовались данные из курсовой за 2010 год («Генотипическое и фенотипическое разнообразие населения Украины по группам крови») (табл. 4). Генетические расстояния определялись по [Nei, 1975]. Генетическое расстояние между двумя популяциями (i и j) рассчитывалось по формуле:
R, r, использовались данные из курсовой за 2010 год («Генотипическое и фенотипическое разнообразие населения Украины по группам крови») (табл. 4). Генетические расстояния определялись по [Nei, 1975]. Генетическое расстояние между двумя популяциями (i и j) рассчитывалось по формуле:
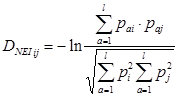
где pai - частота аллеля a в популяции i, l - общее число аллелей в локусах, по которым идет сравнение [2].
Расчет генетических расстояний проводился в программе GDist (ЛПГЧ МГНЦ РАМН).
Смотрите также
Гигиена человека
...
Возможность использования украинских фамилий в качестве квазигенетических маркёров
ВВЕДЕНИЕ
Актуальность
темы. В последнее время использование фамилий в качестве квазигенетических
маркёров стало очень актуальным в решении многих генетических вопросов [7].
Фамилии п ...
Закономерности индивидуального развития. Особенности онтогенеза человека
...