Пигментогенез и формирование различных окрасов у кошкиУчебные материалы по биологии / Генетика окрасов кошек британской породы / Пигментогенез и формирование различных окрасов у кошкиСтраница 1
Палитру окрасов кошек составляют: черный, голубой, шоколадный, коричневый, коричный, лиловый, бежевый, красный, кремовый и желтый цвета. Белый цвет - это отсутствие пигмента (Шустрова, 1997). Пигментов у кошек насчитывается относительно немного (см. раздел 1.1.1). Всё множество расцветок окрасов кошек определяется генами окраски, влияющими на процесс формирования окраса - пигментогенез (Шустрова, 1997).
Пигментогенез начинается в раннем эмбриогенезе. Закладка меланобластов (будущих пигментных клеток) происходит у зародыша в области нервной трубки (Шустрова, 1997). Дифференцировка меланобластов начинается с изменения их формы. Они принимают веретенообразную форму, пригодную для миграции. Они мигрируют сначала в центры пигментации (могут быть расположены в виде пятен на темени, у корня хвоста, на спине или холке), а оттуда - в волосяные фолликулы. Этот процесс находится под контролем гена White(W) (ген доминантного белого окраса). Если этот ген представлен у кошки двумя нормальными рецессивными аллелями w, клетки-предшественницы приобретают нужную для миграции форму. Но если даже один из аллелей является доминантным и мутантным W - клетки теряют способность к организации, остаются на прежнем месте и в будущем пигмента производить не будут, а кошка - носительница доминантного аллеля W останется неокрашенной (Шустрова, 1997).
Ген W может нарушать также формирование близлежащих клеток радужки глаза и кортиева органа. В итоге образуются голубоглазые и даже глухие кошки. В исследованиях Роя Робинсона было показано, что процент голубоглазых кошек среди гетерозигот Ww незначительно ниже, чем среди гомозигот WW. Также на экспрессивность гена W влияют гены модификаторы и регуляторные элементы генома (Сингер, Берг, 1998). В настоящее время известно, что аллель W по признаку собственно белого окрасам показывает почти полную экспрессивность и 100% пенетрантность, по признаку голубоглазости пенетрантность составляет около 40%, по глухоте - около 20% (Шустрова, 1997).
Для того чтобы меланобласты превратились в меланоциты, они должны проникнуть в волосяной фолликул до его окончательного формирования (Рис. 2). Этот процесс определяет насколько окрашенной будет кошка и сохранятся ли у нее белые пятна (Шустрова, 1997).
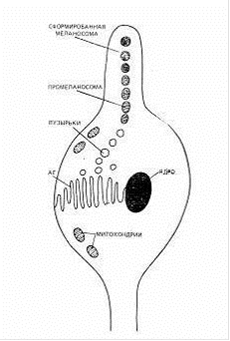
Рис. 2. Строение меланоцита и формирование меланосом
Равная роль в этом процессе отводится гену белой пятнистости (пегости) - Spot
, или, Piebald Spotting
(S). У кошек, гомозиготных по его рецессивному аллелю - ss - окраска развивается полностью. А действие полудоминантного аллеля S вызывает ту или иную степень белой пятнистости. Спектр пегости очень широк: от почти белого окраса с цветным хвостом и пятнами на темени (ванский окрас) до почти полностью развитого окраса с маленьким белым, «медальоном» на груди (Шустрова, 1997).
В последнее время часть фелинологов пришла к выводу, что у гена белой пегости больше двух аллельных состояний (Шустрова, 1997). Предполагают, что ванский окрас определяется наиболее доминантным аллелем серии Sw, а арлекиновый окрас (цветные пятна разбросаны отдельными участками по голове, холке, спине и крестцу) - вторым по степени доминантности аллелем Sp (Шустрова, 1997). Отношения внутри пар этих аллелей не являются жестко определенными, на них также сильно влияют гены-модификаторы (Шустрова, 1997).
Процесс превращения меланобластов в меланоциты контролируется отдельной группой генов. Для превращения аминокислоты тирозина в промеланин, из которого образуется меланин, необходим фермент тирозиназа (Рис. 10). А образование этого регуляторного белка-фермента определяется геном, так называемого локуса Colour
(C). Этот локус представлен целой серией аллелей. Доминантный аллель С обеспечивает синтез нормальной тирозиназы, и тогда особь окрашивается полностью. Рецессивный аллель cs производит термозависимую тирозиназу. Этот фермент активизируется только на холоде. Поэтому гомозиготные по аллелю cs сиамские коты имеют интенсивную окраску только наиболее охлаждаемых, выступающих участков тела - мордочки, ушей, хвоста и лапок. Глаза у них голубые (Шустрова, 1997).
Сходная картина развивается при гомозиготности кошки по другому аллелю серии - cb. Бурманский окрас сохраняет ту же закономерность интенсивного окрашивания выступающих участков тела. При этом глаза золотистые. Гетерозиготы cbcs - так называемые тонкинезы - имеют промежуточный между сиамским и бурманским окрас и бирюзовый цвет глаз. (Шустрова, 1997).
Смотрите также
Почва как среда обитания микроорганизмов
Введение
Микроорганизмы
распространены повсеместно. Весь земной шар "укутан" в живую пленку,
большая доля в которой приходится на микробы. Нет места на нашей планете, ...
Физиология промежуточного мозга. Психофизиология речи и мыслительной деятельности
Введение
Организм
находится в непрерывном взаимодействии с окружающей его внешней средой. Это
взаимодействие очень многогранно; оно обусловлено, с одной стороны, степенью сложности
о ...
Эффекторы
Введение
Система водоснабжения - это комплекс инженерных сооружений,
предназначенных для забора воды из источника водоснабжения, очистки, хранения и
подачи ее потребителю по разводяще ...